Você conhece a diferença entre gramíneas C3 e C4?
Postado em: 11/02/2022 | 8 min de leitura
Escrito por:
Por meio da fotossíntese, as plantas superiores em geral, e até mesmo algas e alguns tipos de bactérias, convertem a energia física da luz solar em energia química. Este processo é essencial para a manutenção de todas as formas de vida.
Desse modo, a fotossíntese pode ser definida como um processo físico-químico, mediante o qual os organismos fotossintéticos sintetizam compostos orgânicos a partir de matéria-prima inorgânica, na presença de luz solar. A fotossíntese é um processo bastante complexo podendo ser analisado em duas etapas: uma etapa fotoquímica, com a presença obrigatória de luz, também chamada de fase clara, e uma segunda etapa, bioquímica ou ciclo fotossintético de redução do carbono, diferenciada segundo o grupo fotossintético ao qual a planta pertence.
A captação da luz é efetuada pelo Complexo de Captação de Luz (CCL) que consiste de uma proteína à qual as clorofilas se associam formando uma espécie de antena que maximiza a energia absorvida. Através de um processo de ressonância, várias moléculas de clorofila transferem a energia para um centro de reação que desestabiliza eletronicamente as clorofilas. Moléculas de água são quebradas e os elétrons dos átomos de hidrogênio são utilizados para o processo de transporte de elétrons.
Os CCL estão acoplados a uma série de proteínas de membrana presentes nos tilacóides, que ao transferirem elétrons acabam reduzindo (adicionando prótons) o NADP transformando-o em NADPH. ATP é também formado durante este processo. A energia contida neste ATP e no NADPH formados será usada no ciclo de Calvin.
Este consiste de um ciclo semi-aberto de reações químicas que executa uma série de adições de moléculas de carbono, utilizando o C do CO2 para alongar moléculas de 3 carbonos (ácido 3 fosfoglicérico – 3PGA) até moléculas de 6 carbonos (frutose-6-fosfato, um açúcar). A maior parte da frutose-6-fosfato formada será utilizada no metabolismo para a produção de amido no cloroplasto e uma outra parte será descarboxilada (isto é, perderá o equivalente a uma molécula de CO2), formando novamente o composto Ribulose 1,5 bisfosfato (RuBP), que entrará no ciclo de carboxilação novamente e reiniciará o ciclo. É por isto que o ciclo de Calvin é considerado semi-aberto.
O CO2 penetra no mesofilo através dos estômatos, sendo, portanto, essencial que estes estejam abertos. A partir da câmara subestomática, o CO2 entra nas células é capturado por um complexo enzimático altamente sofisticado chamado de Ribulose 1,5 bisfosfato carboxilase (RUBISCO) que fica nas membranas dos tilacóides. Este complexo enzimático tem dois substratos, o CO2 e a RuBP, sendo portanto o elemento que fecha o ciclo de carboxilação.
A rubisco não tem apenas dois substratos, mas três. Sabe-se que a rubisco tem também afinidade pelo oxigênio molecular (O2) e as reações associadas à absorção do oxigênio molecular pelas folhas é chamada de fotorrespiração. Ainda não se compreende muito bem as funções fisiológicas da fotorrespiração, mas se sabe que em certas condições, este pode ser um problema sério. Isto porque a concentração de O2 na atmosfera terrestre é de 21% enquanto que a de CO2 é de apenas 0,038%, o que aumenta grandemente a probabilidade da rubisco capturar O2 ao invés de CO2. Com isto, a planta tem que lidar com uma assimilação aparentemente indesejável de O2. Indesejável porque ao entrar na planta o O2 tem o potencial de formar radicais livres que são altamente deletérios para o metabolismo. Desta forma, o sistema tem que gastar uma quantidade razoável de energia e nutrientes (O custo da fotorrespiração para a fotossíntese do tipo C3 está entre 30 a 40%) para evitarem danos sérios, livrando-se do O2. Por este raciocínio, a fotorrespiração seria um “peso” para o metabolismo C3, roubando energia que poderia ser usada para o crescimento.
Como se observa na Figura 1, há dois caminhos de carboxilação. Em um deles a via tem início com um composto de 3 carbonos (3PGA) e por isso é chamada de via C3. Plantas como soja, milho feijão e todas as árvores que tiveram a fotossíntese estudada até o momento, usam a via C3 para a fotossíntese.

Algumas plantas, principalmente gramíneas (cana-de-açúcar e milho p.ex.) e parte das bromélias, desenvolveram um sistema complementar à via C3 chamado de via C4. Este sistema permite à folha o armazenamento de ácidos com 4 carbonos antes de estes serem captados pela rubisco. Neste caso há uma mudança morfológica importante que é a existência de uma bainha vascular, uma camada adicional de células que envolve os feixes vasculares. O CO2 é captado nestas células do mesofilo pela enzima Fosfo Enol Piruvato carboxilase (PEPc), presente nas células do mesofilo, a qual forma um composto de 4 carbonos que poderá ser descarboxilado a 3PGA e usado pela rubisco, presente nas células da bainha vascular. Nas células da bainha vascular, o ácido (malato por exemplo) e descarboxilado, formando CO2 novamente. Este mecanismo causa um aumento espetacular na concentração de CO2 na célula da bainha em relação à do mesofilo. Enquanto a pressão parcial de CO2 no mesofilo é da ordem de 150 µbar, na bainha vascular chega ser dez vezes maior (1500 µbar). Com isto, a rubisco fica em uma situação em que a concentração de substrato é muito alta, evitando a competição do oxigênio que leva à fotorrespiração.
=> Gosta do conteúdo? Você também pode gostar do conteúdo completo do curso Planejamento forrageiro para a seca. O curso pode ser adquirido individualmente ou você pode optar por assinar a plataforma EducaPoint, tendo acesso a todos os cursos disponíveis (mais de 245!) por um preço único. Clique aqui para assinar.
=> Gosta do conteúdo? Você também pode gostar do conteúdo completo do curso Planejamento forrageiro para a seca. O curso pode ser adquirido individualmente ou você pode optar por assinar a plataforma EducaPoint, tendo acesso a todos os cursos disponíveis (mais de 245!) por um preço único. Clique aqui para assinar.
Como um dos grandes problemas das plantas é a perda de água pelos estômatos quando estes estão abertos para permitir a entrada do CO2, o mecanismo C4, ao aumentar em dez vezes a concentração deste gás nas células da bainha vascular, acaba evitando a perda de água, pois o aproveitamento do CO2 é muito melhor do que em plantas C3.
A existência da via C4 é um exemplo interessante da relação estrutura-função em plantas. A bainha vascular e seus mecanismos bioquímicos acoplados à via C3 criaram, durante a evolução, uma espécie de “bomba” que torna o sistema fotossintético mais eficiente em certas situações, praticamente eliminando a fotorrespiração.
Há várias consequências importantes da presença da bainha vascular. Uma delas é que as taxas de absorção de CO2 são muito mais altas, pois o sistema pode armazenar mais carbono de forma intermediária (no ácido C4) tornando a planta relativamente menos dependente de controlar a abertura e fechamento de estômatos para prevenir a perda de água.
O resultado é que quando se compara o rendimento quântico de plantas C3 e C4 em diferentes temperaturas (Figura 2), nota-se que as diferenças entre o desempenho dos dois sistemas em diferentes temperaturas tem vantagens e desvantagens que dependerão do clima onde vivem. Enquanto as plantas C4 tem desempenho constante em temperaturas que variam entre 10 e 40o C, as C3 apresentam uma queda linear em desempenho quando se aumenta a temperatura. É instrutivo analisar as duas em um mesmo gráfico (Figura 2).
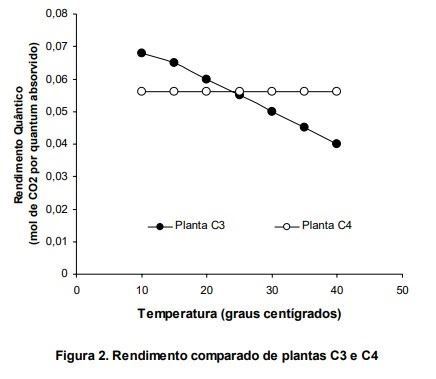
Veja que as C3 levam vantagem até cerca de 28o C, mas que, devido ao fato das C4 não apresentarem queda de rendimento em temperaturas mais altas, estas últimas passam a ter maior eficiência relativa acima de 28o C. O fato de as plantas C4 lidarem melhor com temperaturas mais altas também permitem que seus sistemas de captação de luz possam suportar intensidades luminosas muito maiores. Com isto, enquanto as plantas C3 funcionam bem entre 400 a 500 µmoles de fótons.m-2.s-1, as plantas C4 podem funcionar em intensidades maiores do que 2000 µmoles de fótons.m-2.s-1.
Se considerarmos as intensidades luminosas normalmente encontradas em condições naturais, fica claro que plantas C3 como as árvores tropicais por exemplo, se adaptam bem a condições mais sombreadas (veja a diferença entre os pontos de compensação de C3 e C4 na Tabela 1), enquanto as gramíneas C4 ou bromélias que crescem em campos rupestres conseguem se desenvolver melhor em altas intensidades luminosas e em altas temperaturas.
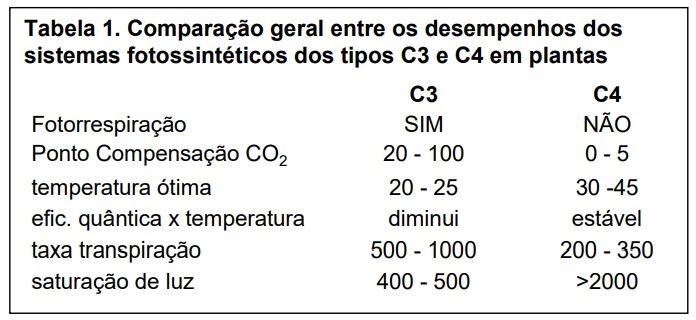
Uma outra questão importante é como os dois tipos de planta lidam com a água. Como pode ser visto na Tabela 1, as plantas C4 são bem mais eficientes que as C3 em lidar com a água. Como mencionado acima, isto se deve à maior eficiência em captar e armazenar o carbono oriundo do CO2, isto é, a bomba de armazenamento do ácido de quatro carbonos, o que permite às plantas C4 um gerenciamento melhor da abertura estomática, que é um processo fundamental no controle da transpiração foliar.
Tais características são extremamente importantes se considerarmos, por exemplo, uma gramínea em um campo cerrado vivendo em temperaturas altas e em baixa umidade durante o dia. Por isto, as gramíneas C4 tendem a se beneficiar de suas vantagens em relação às C3 em condições extremas, para invadir regiões mais áridas do planeta como os desertos australianos, as savanas africanas e americanas e os campos de gramíneas do noroeste da América do Norte.
Note que como o clima do planeta varia em escala geológica, as regiões em que existirão condições propícias para os diferentes tipos de plantas variarão também. Estes ciclos aliados às alterações relativamente simples para transformar o sistema C3 em C4, parecem estar associados ao fato de que, durante a evolução, este último apareceu várias vezes (acredita-se que pelo menos 45 vezes) de forma independente em vários grupos pouco relacionados filogeneticamente.
* Baseado no texto do professor Marcos S. Buckeridge, Comparação entre os sistemas fotossintéticos C3 e C4.
Mais informações:
contato@educapoint.com.br
Telefone: (19) 3432-2199
WhatsApp (19) 99817- 4082
Fonte consultada:
Comparação entre os sistemas fotossintéticos C3 e C4, por professor Marcos S. Buckeridge, e monitores Adriana Yepes, Amanda Pereira de Souza, Mauro Marabesi, Patrícia Pinho Tonini (http://saulcarvalho.com.br/wp-content/uploads/2016/10/sistemas-c3-c4.pdf)
Acesse os conteúdos relacionados a esse post: